

Evaluación del estrés hídrico y de tres índices de vegetación en la uva de vino, Vitis vinifera, en el Valle de Guadalupe, Baja California, México
Assessment of Plant Water Stress and Three Vegetation Indices in the Grapevine, Vitis vinifera, from the Guadalupe Valley, Baja California, Mexico
Rodrigo Méndez Alonzo, Ramiro Santos Cobos, Anaid Rodríguez Bastarrachea, Yadira Romero Toledo, Samantha Díaz de León Guerrero, Eulogio López y Mario Salazar*
Nota: el presente estudio fue financiado por medio del Fondo Sectorial CONACYT-INEGI proyecto 278755-2016-1; agradecemos el apoyo tanto técnico durante este proyecto de los biólogos Amos Antonio Pérez Hernández y Arturo Victoria Hernández✟ (INEGI) como administrativo del personal del Centro de Investigación Científica y de Educación Superior de Ensenada (CICESE); Ramiro Santos, Anaid Rodríguez y Samantha Díaz de León agradecen al Posgrado de Ciencias de la Vida CICESE el apoyo para la realización de sus tesis y al CONACYT por las becas de posgrado otorgadas; asimismo, los autores agradecen al personal del viñedo Monte Xanic las facilidades prestadas para este estudio.
* CICESE, mendezal@cicese.mx, ramirosnts@gmail.com, anaidbastarrachea@gmail.com, yromerotoledo@gmail.com, dsamantha@cicese.edu.mx, elopez@cicese.mx y msalazar@cicese.mx, respectivamente.
Vol. 12, Núm. 2 EPUB Evaluación del estrés hídrico... EPUB

|
El potencial hídrico en plantas (Ψ, una medida de disponibilidad hídrica) varía en función del volumen de irrigación, especialmente en zonas áridas y semiáridas. En el presente estudio probamos si menores volúmenes de irrigación producirían valores menores del Ψ y de los índices de vegetación NDVI, PRI y WBI en Vitis vinifera, la uva de vino, en el Valle de Guadalupe, Baja California, México. En noches de verano, con irrigación, el Ψ fue de -0.3 MPa, mientras que a medio día llegaba a -2.5 MPa. En otoño, sin riego, el Ψ fue de -1 MPa durante la noche y -2.0 MPa en el día. El NDVI mostró valores entre 0.56 durante mayo a 0.23 durante diciembre, en hojas senescentes. El Ψ nocturno varió más en concordancia con la hipótesis, lo que sugiere ventajas de la irrigación vespertina o nocturna en la uva en el Valle de Guadalupe. Palabras clave: agricultura sustentable; irrigación; potencial hídrico; viticultura.
|
Plant water potential (Ψ, a measure of water availability) varies as a function of irrigation volume, particularly in arid and semiarid zones. In the present study, we tested wether lower irrigation volumes would produce lower values of Ψ as well as of three vegetation indices (NDVI, PRI, and WBI) in the grapevine, Vitis vinifera, in the Guadalupe Valley, Baja California, Mexico. On summer nights, with irrigation, Ψ reached -0.3 MPa, while at midday it reached -2.5 MPa. In autumn, without irrigation, Ψ was -1 MPa at night and -2.0 MPa during the day. NDVI values ranged from 0.56 in May to 0.23 throughout December, in senescent leaves. Nocturnal Ψ varied more in agreement with the hypothesis, suggesting advantages of evening or night irrigation on grapes in the Guadalupe Valley. Key words: irrigation; plant water potential; sustainable agriculture; viticulture. |
Recibido: 12 de diciembre de 2019.
Aceptado: 16 de marzo de 2021.
Introducción
Los eventos meteorológicos extremos, como la sequía, se han vuelto más recurrentes y de mayor intensidad a consecuencia del cambio climático producido por el incremento de las emisiones de gases de efecto invernadero y la reducción en extensión de los ecosistemas que permiten la captura y el secuestro de carbono (IPCC, 2018). Es un evento que puede considerarse desde distintos puntos de vista: A. Dai clasificó a los tipos de sequía en hidrológica, que es aquella donde los niveles de los reservorios (ríos, lagos y freáticos) están de manera persistente debajo de los valores promedios desde su registro; meteorológica, que se define por la precipitación anormalmente baja para una región en comparación con el registro histórico; y agronómica, que resulta de la combinación de precipitación baja y evapotranspiración alta que no permite el mantenimiento de la productividad agrícola (Dai, 2011 y 2013; Trenberth et al., 2014). A estas clasificaciones es posible agregar la definición de sequía desde el punto de vista de la fisiología de plantas, donde el estrés hídrico se considera como la reducción en la conductividad hidráulica de la planta a consecuencia de la reducción en el contenido de agua en el suelo (Tyree y Ewers, 1991; Cochard, 2014).
El transporte de agua en las plantas ocurre de manera continua y, por lo general, unidireccional a través de una corriente que transcurre en dirección suelo-raíz-tallo-hoja-atmósfera (Sperry et al., 2003). Este continuo de transportación hidráulica es consecuencia de la interacción entre fuerzas moleculares no covalentes que cohesionan a las moléculas de agua entre sí y que, además, causan su adhesión con conductos llamados vasos (o traqueidas en coníferas y otras plantas) dentro de la madera de estas (técnicamente llamada xilema). La fuerza que genera el impulso inicial a este flujo es la desecación ocasionada por la liberación continua de agua en cámaras especializadas en el intercambio de gases en el mesófilo de las hojas (Nobel, 1999; Taiz y Zeiger, 2010).
Dentro de las hojas, en las estructuras conocidas como criptas estomáticas foliares, ocurre la liberación de agua a la atmósfera y la absorción de moléculas de CO2, ocasionando un gradiente de deshidratación que causa tensión en una columna continua de agua, lo cual impulsa a que se mueva desde la frontera de la raíz con el suelo hasta las criptas estomáticas, donde ocurre la transpiración (Evert, 2006). Cuando el nivel de saturación relativa de agua en el aire circundante (déficit de presión de vapor) disminuye, incrementando la intensidad de la desecación, aumenta la transpiración de las plantas, que depende, a su vez, del área foliar, densidad y tamaño de estomas y conductancia de la red de venas en las hojas (Nobel, 1999).
Este mecanismo de transporte se conoce como teoría de tensión-cohesión y es susceptible de ser cuantificado mediante el potencial hídrico (Ψ), que es la comparación de la energía libre de cualquier solución respecto al agua pura (Dixon y Joly, 1895; Scholander et al., 1965; Tyree y Ewers, 1991; Tyree y Zimmermann, 2013).
El Ψ refleja la capacidad de movimiento del solvente de una solución de un compartimento a otro, y en plantas puede ser utilizado para comparar la cantidad de agua susceptible de ser movilizada de uno con mayor Ψ (Ψ = 0 para el agua pura) hacia otro de menor Ψ a través de un gradiente de concentración de agua (Boyer, 1995; Nobel, 1999). Esta analogía de potenciales hidráulicos con respecto a sistemas eléctricos y químicos permite hacer comparaciones relativas del estado de estrés hídrico experimentado por diferentes especies de plantas, o bien, variedades o tratamientos experimentales dentro de una misma especie. Cuando estas mantienen la transpiración a través de las hojas, pero tienen el Ψ que no permite la extracción de agua del suelo (Ψplanta > Ψsuelo), el tejido foliar comienza a colapsar y los vasos del xilema empiezan a presentar embolismos, que son burbujas de aire que bloquean el flujo, reduciendo la conductividad hidráulica del tallo o de la hoja (Sperry y Tyree, 1988; Tyree y Sperry, 1989). Las plantas pueden, en tal caso, evitar la pérdida de agua mediante el cierre estomático, pero si la sequía es persistente y la disfunción hidráulica es sostenida en el tiempo, agotan sus reservas de carbono y son más susceptibles al ataque de patógenos. Es en estos puntos que puede ocurrir la pérdida de productividad y, finalmente, la mortalidad, y es la razón por la cual es necesaria la irrigación para mantener el estado hídrico favorable de las plantas (McDowell et al., 2008; McDowell, 2011).
Una de las ventajas de cuantificar el Ψ en plantas es que permite ayudar a establecer calendarios de irrigación en especies leñosas (Shackel et al., 1997; Améglio et al., 1999; Patakas et al., 2005). Una vez realizado el riego, el agua puede tener tres destinos: 1) movilizarse al manto freático a través de la percolación de esta por el suelo o por movimiento horizontal a través del gradiente gravitacional, 2) evaporarse directamente de los poros del suelo a la atmósfera y 3) ser atrapada en micorrizas, pelos radiculares y raíces leñosas de las plantas para su traslado al sistema vascular y su uso posterior en transpiración y en sus funciones vitales (Boyer, 1995). Conocer la variación en el Ψ en diferentes cultivos permite establecer valores de referencia dentro de los cuales las plantas mantienen correctamente su función hidráulica, tanto diaria como estacional (Patakas et al., 2005), permitiendo reducir las pérdidas por percolación o evaporación.
Por otra parte, los valores del Ψ generalmente están asociados a conjuntos de características fenotípicas de las plantas, como la densidad de madera y hojas, el área foliar, la fenología foliar, entre otras (Jacobsen et al., 2008). El conocimiento de los valores de referencia del Ψ y de otras características estructurales y funcionales puede coadyuvar al establecimiento de lineamientos de riego mínimos y máximos diarios y estacionales, mejorando las decisiones de manejo de irrigación en diferentes cultivos, complementarios a los valores de evapotranspiración calculados a partir de métodos meteorológicos.
Por otra parte, la estacionalidad y variabilidad en disponibilidad de recursos (principalmente luz, agua y nutrientes) modifican las características espectrales foliares, en particular en ambientes áridos y estacionales. Las firmas espectrales en hojas se deben a la composición de pigmentos, en especial clorofilas y pigmentos accesorios, y es variable a escalas inter e intraespecíficas (Carter y Knapp 2001; Cavender-Bares et al., 2017; Meireles et al., 2020).
Dado que los pigmentos están íntegramente relacionados con la función fisiológica de las hojas, en particular con la asimilación de carbono (Gamon et al., 1997), la estimación de índices de vegetación a partir de las firmas espectrales de hojas es una herramienta que provee información sobre el desempeño de las plantas, pudiendo cuantificarse a escala de la hoja o mediante sensores remotos (Blackburn, 1998). Los índices de vegetación son medidas cuantitativas, producto de varios valores espectrales que son sumados, divididos o multiplicados en ecuaciones informativas respecto al estado fisiológico de la vegetación monitoreada (Myneni et al., 1995). Su uso es particularmente relevante en los ecosistemas semiáridos, donde la disponibilidad hídrica varía de manera radical a lo largo del año (Elmore et al., 2000; Schmit y Karnieli, 2000). En este tipo de ambientes se puede esperar que los espectros de absorción y reflexión de la luz, que dependen de la concentración de pigmentos en la lámina foliar, se asocien al uso de agua, pues la absorción de luz en el infrarrojo cercano está fuertemente influenciada por la cantidad de agua presente en el tejido foliar (Pettorelli et al., 2005). Por lo tanto, el utilizar conjuntos de datos fisiológicos y su relación con los índices de vegetación, como el de Verdor (NDVI, Tucker, 1979), los fisiológicos (PRI, Garbulsky et al., 2008) y con los de agua (WBI, Claudio et al., 2006), puede dar pistas sobre el desempeño de las plantas, tanto de manera remota como medidas in situ (Pettorelli et al., 2005).
Conocer la variabilidad en el Ψ y en valores de índices de vegetación permite la cuantificación del estado fisiológico de las plantas, lo que puede contribuir a mejorar el manejo de irrigación en regiones áridas y semiáridas (en las cuales el potencial de evapotranspiración es mayor que la precipitación acumulada anual). Contar con valores de referencia del Ψ y de índices de vegetación medidos in planta es un paso necesario para la validación del estrés hídrico mediante sensores remotos, a través de diferentes temporadas del año y bajo diversas condiciones de irrigación.
El presente estudio tuvo como objetivos caracterizar la variabilidad del Ψ en hojas y tallos a lo largo de experimentos estacionales y diarios, así como la variabilidad estacional de tres índices de vegetación en hojas de Vitis vinifera, L, la uva de vino. El Valle de Guadalupe (VG) es la zona vitivinícola más importante del país, bajo condiciones ambientales semiáridas (298 mm por año, Del Toro et al., 2019), lo cual implica que es necesario proporcionar mejor información sobre los requerimientos hidráulicos de los cultivos. Nuestra hipótesis supone que la variación estacional y menores volúmenes de irrigación deberían producir valores más negativos en el Ψ y menores en los índices NDVI, PRI y WBI. Previo a esta prueba de hipótesis, generamos una caracterización de las condiciones edáficas en el VG y cuantificamos 16 métricas estructurales y fisiológicas del tallo y hoja. En conjunto, estas mediciones proveen información para mejorar la precisión en irrigación monitoreada por variables fisiológicas en los cultivos de uva del VG, México.
Metodología
Área de estudio
El Valle de Guadalupe se localiza en el extremo noroeste del estado de Baja California (BC), México. Es un valle intermontano de cerca de 94 km2 localizado al noreste de la ciudad de Ensenada, BC. Se ubica entre los paralelos 32°08’-32°00’ N y 116°41’-116°29’ W. Tiene una elevación que varía entre los 300 y los 370 metros sobre el nivel del mar (m s. n. m.), con pendientes suaves (menores a 10°), y se encuentra rodeado por laderas empinadas (entre 40-60°), que se elevan de los 400 a los 750 m s. n. m. La precipitación media anual promedio es de 300 mm, de los cuales cerca de 80 % ocurre en invierno, en los meses de diciembre a marzo. La temperatura media anual es de cerca de 18 °C, variando de los 13 a los 24 °C (Del-Toro et al., 2019). El VG forma parte de la extensión sur de la Provincia Florística de California, un área de alto endemismo en diversidad biológica, caracterizado por un clima mediterráneo, con inviernos frescos y lluviosos y veranos secos y cálidos (Rundel et al. 2016).
Debido a sus características climáticas y edafológicas, las actividades productivas en el Valle de Guadalupe son eminentemente agrícolas (Espejel et al., 1999); 60 % del área cultivada (cerca de 4 mil hectáreas) se destina a la uva de vino (Vitis vinifera L.) y el restante, al olivo (Olea europaea L.), algarrobo (Ceratonia siliqua L.), cítricos y otros frutales (Secretaría de Fomento Agropecuario, BC, 2015). Estos cultivos se riegan, principalmente, con agua de los freáticos, pues los cuerpos de agua superficiales son virtualmente inexistentes (Espejel et al., 1999; Daesslé et al., 2006). El VG es la región vitivinícola más importante en el país, acumulando entre 90 y 95 % de la producción de vinos mexicanos. El cultivo comercial de uva en esta región data de la década de los 30 del siglo pasado, cuando el presidente Abelardo L. Rodríguez fomentaba la producción de olivos y vides en el Valle de Guadalupe. Noventa años después, el Valle de Guadalupe cuenta con cientos de casas vitivinícolas y se ha convertido en un polo de atracción turística a nivel estatal (Espejel et al. 1999).
Determinación de propiedades físico-químicas del suelo en el VG
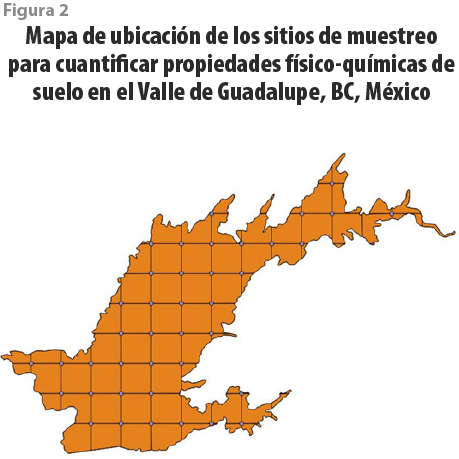
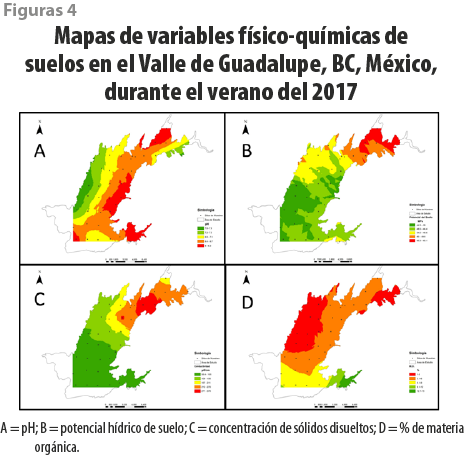
Para poder entender la heterogeneidad de las propiedades físico-químicas del suelo al interior de la zona agrícola del Valle de Guadalupe, se realizó un mallado, tomando un punto de muestreo cada 2 km, para un total de 38, cubriendo un área aproximada de 60 km2 (ver figura 2). En cada punto, se realizaron excavaciones con una pala y un barreno para suelos hasta una profundidad aproximada de 40 cm; una vez que se extraía tierra entre los 25-35 cm, se les retiraba todo tipo de materia orgánica visible y se agregaba a un recipiente de medio litro para tener control del volumen de suelo que se tomaba por punto. Después, las muestras se guardaban en bolsas de plástico con sello, se etiquetaban y transportaban al laboratorio el mismo día. Estas fueron separadas para las siguientes determinaciones: potencial hídrico del suelo (Ψsuelo), pH, porcentaje de materia orgánica y contenido de sólidos disueltos totales (Romero-Toledo, 2017).
El Ψsuelo se cuantificó por medio de un potenciómetro de punto de rocío (WP4-T, Metter Inc., Pullman, WA, USA), a partir de muestras de 20 ml obtenidas cada mes a una profundidad de 20 cm; el pH y el contenido de sólidos disueltos fue medido en solución (10 ml suelo × 80 ml agua bidestilada) con un potenciómetro de pH (A221Orion Star Portable, Thermo Scientific, Beverly, MA, USA); el porcentaje de materia orgánica fue cuantificado mediante diferencia de masas tras la ignición en muestras de 10 ml (presecado en un horno a 105 °C, Fisher Isotemp, 500 series, Pittsburgh, PA, USA, ignición en una mufla a 280 °C por cuatro horas, 2-525 muffle furnace, J.M. Ney, Tucson, AZ, USA); y, finalmente, para cada punto, se cuantificó la composición de textura de suelo por el método de higrometría (Bouyoucos, 1962).
La distribución espacial en el área agrícola del Valle de Guadalupe de las características físico-químicas del suelo se extrapolaron al espacio no muestreado mediante la herramienta Kriging Bayesiano en ArcGis 10.1 (Environmental Systems Research Institute, Redlands, CA, EUA).
Experimento de variabilidad diaria y estacional en el potencial hídrico
Dentro del VG se seleccionó para estudiar la variación diaria en el Ψ en hojas y tallos al viñedo Monte Xanic (32.0962 N, 116.5900 O), por tener permisos del propietario y por contar con la instalación de una torre de medición de parámetros micrometeorológicos.
Para esta investigación, se utilizaron seis individuos de la variedad Cabernet Franc sobre una misma hilera, cultivados en orientación noroeste a sureste, con edad aproximada de nueve años. Cada planta de estudio estuvo separada de la subsecuente por otra para evitar la posibilidad de muestrear ramas del mismo individuo (densidad de cultivo de ca. 3 mil plantas por hectárea). Se evitaron seleccionar aquellas que se encontraban junto al final de la hilera —tomando en cuenta que la extensión de la línea seleccionada superara las 12 plantas—, la cual estaba en el área de medición de una estación experimental micrometeorológica para la medición de covarianza de turbulencias.
La textura del suelo resultó franco-arenosa, con composición textural de 63 % arenas, 27 % limos y 10 % arcillas; el pH de suelo fue de 6.2; el contenido total de sólidos disueltos, de 357 mS/cm; y el porcentaje de materia orgánica, de 5.1 por ciento.
Durante el verano del 2017, el régimen de riego consistió en dos eventos a la semana, con tres días de intervalo entre uno y otro. Estos se realizaron por goteo, durante seis horas, con gasto de 2 litros por hora (12 por planta, 24 por semana). El riego se suspendió en los primeros días de octubre y se mantuvo sin este durante los meses posteriores en otoño e invierno. En la primavera y el verano del 2018, el riego fue por goteo cada seis días durante aproximadamente siete horas, con gasto de 2 litros por hora.
Se hicieron tres campañas de medición, del 10-13 de agosto de 2017 (postenvero, maduración de bayas), 13-17 de noviembre de 2017 (senescencia foliar) y 7-12 de junio de 2018 (envero, cambio de coloración en bayas). En cada una se realizaron mediciones de forma continua del Ψ cada dos horas durante 72 horas para obtener 36 registros por individuo. Para la medición del Ψ, las ramas fueron introducidas durante 10-15 minutos en una bolsa opaca con papel saturado con agua. El material fue procesado de inmediato tras la toma de muestras cada dos horas con el método de cámara de presión (Scholander et al., 1965), con un equipo modelo 1505D (PMS Instruments, Albany, OR, USA). Simultáneamente, se hicieron mediciones consecutivas de la conductancia de los estomas (gs) en una sola rama por individuo por especie. Las ramas fueron seleccionadas con exposición hacia el sur, independientes de las utilizadas para cuantificar el Ψ. Las mediciones de conductancia se realizaron en la cara abaxial de las hojas con un porómetro foliar (SC-1 Decagon Devices Inc, Pullman, WA, USA). Previo a esto, la humedad o la suciedad de las hojas fue eliminada con papel secante. Para la obtención de medidas precisas, se llevaron a cabo calibraciones del equipo cada seis horas, más o menos, durante el transcurso del día, o a una mayor frecuencia si ocurrían cambios marcados de temperatura (°C) y humedad relativa (%). En cada medición de conductancia se corroboró la temperatura ambiental y humedad relativa a 1 cm aproximadamente de la superficie abaxial de la hoja utilizando un termohigrómetro (Traceable humidity temperature pen, Thomas Scientific).
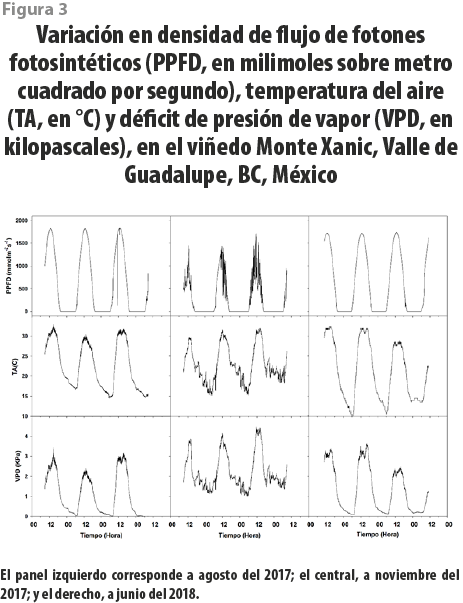
Durante las fechas de medición, se cuantificó la variación en temperatura ambiental (T), déficit de presión de vapor (VPD, calculado a partir de la T y humedad relativa, RH) y la densidad en el flujo de fotones fotosintéticos (PPFD), mediante la extracción de datos de la instalación experimental mencionada anteriormente. En la misma zona, aledaña a las plantas muestreadas, se cuantificó el contenido de agua en el suelo con sensores tanto de temperatura y humedad de suelo (Hydra Probe II, Stevens, Portland, OR, USA) como para medir los flujos de calor de suelo (HFP01, Hukseflux, Delf, The Netherlands). La frecuencia de extracción de datos correspondía a los 10 minutos previos a cada hora de medición y a los 10 minutos posteriores, con una frecuencia de 0.016 Hz (esto es, un conjunto de datos por minuto). La información de cada variable se promedió y su dispersión se evaluó por desviaciones estándar. T y RH fueron cuantificadas mediante un sensor sobre 2 m del suelo (T-RH sensor HMP155, Vaisala, Helsinki, Finland), y PPFD fue cuantificado con un radiómetro a la misma altura (LI 190R, LI-COR, Lincoln, NE, USA) (ver figura 3).
Las trayectorias del Ψ fueron correlacionadas con variables meteorológicas y con gs usando correlaciones lineales de Pearson en el programa JASP 0.8.4.0 (JASP Team Amsterdam).
Determinación de índices de vegetación en hojas
Para cuantificar la variación en patrones espectrales foliares, se realizaron mediciones durante el final de la temporada seca, desde septiembre del 2017 hasta mayo del 2018, teniendo que detener el muestreo por problemas logísticos. Estas se llevaron a cabo en los mismos seis individuos que se utilizaron para medir el potencial hídrico y la conductancia de los estomas. Para cada individuo, se marcó una rama de aproximadamente 25 cm, terminal orientada al sur, que fuera distinta a las utilizadas para cuantificar la gs. Cada mes, se registraron los espectros de absorción y reflexión de la luz en el rango de los 300 a 1 100 nm en una hoja sana y expandida.
Durante el año de medición no hubo eventos completos de defoliación por invierno, por lo cual se pudo dar seguimiento a lo largo del año. Las firmas espectrales fueron registradas mediante un espectro-radiómetro portátil (PP Systems, UNISPEC SC Single Channel, Amesbury, Massachusetts, EUA), con rango de medición entre los 300-1 100 nm. A partir de las firmas espectrales, se extrajeron las bandas correspondientes al infrarrojo cercano (NIR, por sus siglas en inglés, abarcando de los 800-900 nm), rojo (R, de los 600 -650), rojo superior (RED_S, 564-576), rojo inferior (RED_I, 525-537), infrarrojo superior (NIR_S, 954-986) e infrarrojo inferior (NIR_I, 884-916). Con esta información, se calcularon los siguientes índices de vegetación (Tucker, 1979; Peñuelas et al., 1993; Gamon et al., 1997; Pettorelli et al., 2005):
- De Diferencia Normalizada (Normalized Difference Vegetation Index, NDVI), calculado mediante:
NDVI= (NIR - RED) / (NIR + RED).
- De Reflectancia Fotoquímica (Photochemical Reflectance Index, PRI):
PRI= (RED_S - RED_I) / (RED_S + RED_I).
- De la Banda del Agua (Water Band Index, WBI):
WBI= (NIR_I) / (NIR_S).
Estos índices se basaron en los valores encontrados en hojas durante muestreos mensuales realizados entre octubre del 2017 y mayo del 2018. Se asociaron los valores de los índices utilizados con los del Ψ y de gs mediante correlaciones lineales de Pearson en el programa JASP 0.8.4.0 (JASP Team Amsterdam).
Características estructurales y funcionales de la vid en el VG
De las mismas seis plantas a las cuales se midió el Ψ y la conductividad de los estomas, se obtuvieron un mínimo de cinco hojas por cada individuo para la cuantificación de características funcionales (área foliar, masa foliar por unidad de área, contenido de masa seca en hoja y densidad foliar) y del tallo (porcentaje de masa seca en tallo, cinco tallos) (Pérez-Harguindenguy et al., 2013). Además, se realizaron cuantificaciones anatómicas del área de tejido en tallo, en segmentos de 1 cm de diámetro cortados transversalmente. Estos cortes fueron examinados con un microscopio estereoscópico (NIKON C-LEDS, Melville, NY, EUA) para obtener las proporciones de área ocupadas por la corteza, el xilema y la médula.
Como complemento a la determinación de los valores del Ψ, se calculó el potencial hídrico en el punto de marchitamiento (Ψtlp). Esta medida es un indicador de la concentración de solutos intracelulares en hojas, donde el mayor valor determina un Ψ más negativo (Bartlett et al., 2012a), el cual se estimó mediante la extracción de una muestra de 1 cm2 de tejido foliar para una hoja de cada uno de los seis individuos muestreados. Estas muestras se congelaron en nitrógeno líquido por, al menos, dos minutos. Posteriormente, el Ψtlp se cuantificó en un osmómetro de vapor siguiendo los métodos de Bartlett et al. (2012b).
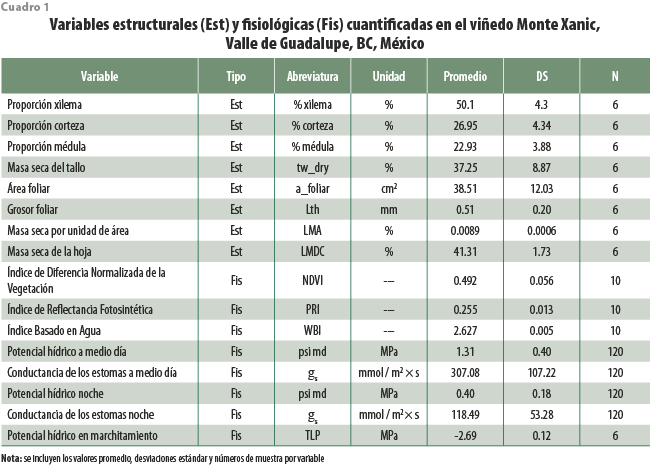
Dado que las mediciones de estas variables no ocurrieron en la misma frecuencia que las del potencial hídrico, conductancia de los estomas y de índices de vegetación, se presentan los datos de estadística descriptiva para cada variable sin generación de pruebas de hipótesis. Las 16 características anatómicas, fisiológicas y ecológicas de la planta de vid —abarcando caracteres de tallo, hoja, espectrales de hoja y de arquitectura hidráulica del tallo (ver cuadro 1)— se obtuvieron de las mismas plantas que fueron estudiadas para la cuantificación del Ψ, gs y espectroscopía foliar.
Resultados
Propiedades físico-químicas del suelo del Valle de Guadalupe
Encontramos que los suelos de mayor acidez (valores entre 5.8 hasta 7.73, figura 4A) y concentración de materia orgánica (de 1.3 hasta 12.8 %, figura 4D) están ubicados en la región sur del Valle de Guadalupe. Los valores del potencial hídrico del suelo fueron variables en la mayor parte del Valle (figura 4B). La concentración de solutos disueltos en agua es mayor en el noreste del VG y menor en el sur (valores entre 25 hasta 749 mS / L, figura 4C).
Trayectoria diaria y estacional del potencial hídrico y conductancia de los estomas
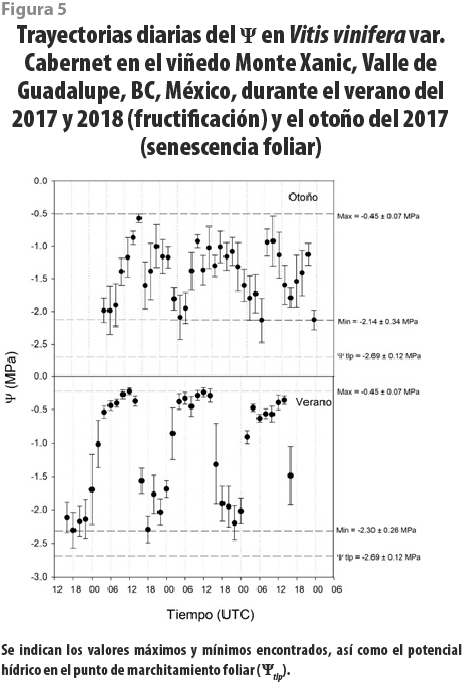
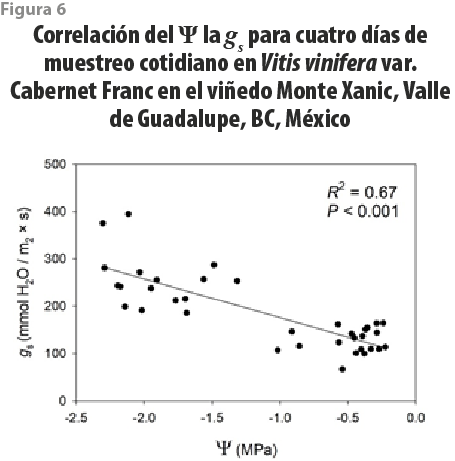
Se realizó un seguimiento cotidiano del Ψ y la gs en tres fechas (agosto del 2017, noviembre del 2017, junio del 2018) para evaluar los efectos de los cambios en los regímenes de irrigación en el viñedo Monte Xanic. Los valores máximos del Ψ durante el verano fueron de -0.45 MPa, ocurriendo tras caer la noche, mientras que los mínimos, de -2.5 MPa a medio día. Por otra parte, en otoño, los máximos del Ψ durante la noche fueron de -0.45 MPa, aunque la moda estuvo en -1 MPa, y los mínimos, de -2.1 MPa. En ningún caso, los valores mínimos llegaron al punto de marchitamiento (ver figura 5). Los de gs para medio día en el verano fueron de 224 ± 74 mmol / m2 × s, mientras que, para la noche, fueron de 132 ± 21 mmol / m2 × s. En otoño, los valores diurnos y nocturnos de gs fueron de 220 ± 48 mmol / m2 × s y de 122 ± 16 mmol / m2 × s. Se encontró que los patrones diarios del Ψ y de gs estuvieron positivamente correlacionados (r2 = 0.67, ver figura 6).
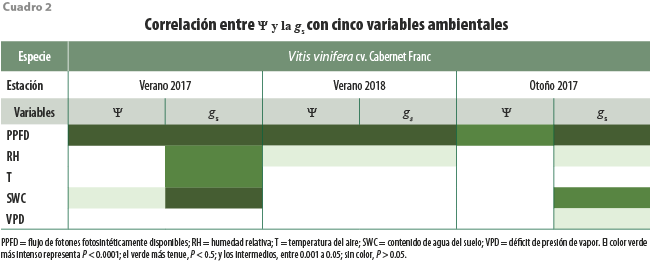
Tanto los valores del Ψ como de gs estuvieron asociados en alguna fecha con todas las variables ambientales cuantificadas. Sin embargo, la única que estuvo asociada de manera consistente con estas variables fisiológicas fue el flujo de fotones fotosintéticamente disponibles (ver cuadro 2).
Trayectoria de variación espectral en hojas
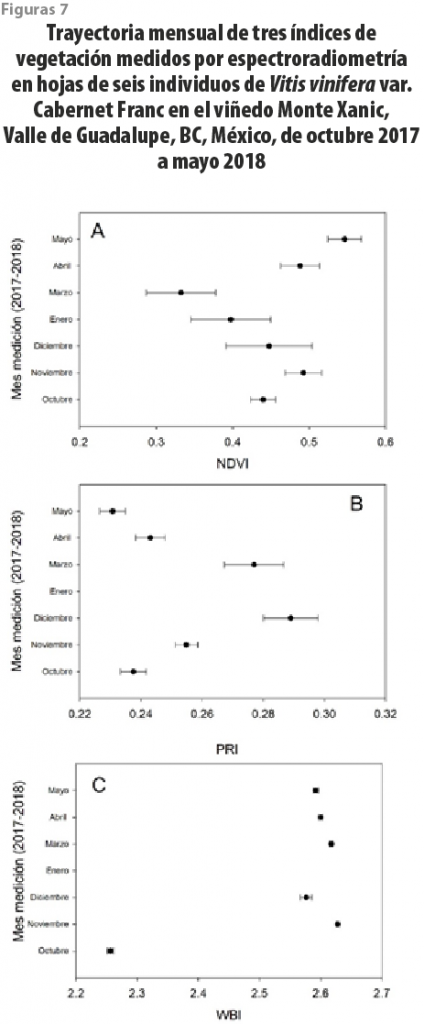
Los valores de los tres índices fueron altamente variables, encontrándose para el NDVI de 0.44 ± 0.017 (error estándar) para octubre del 2017, y reduciéndose por senescencia hasta 0.33 ± 0.045 en marzo. Al mismo tiempo, en hojas nuevas se registraron datos de 0.39 ± 0.052 en diciembre, hasta 0.55 ± 0.022 en mayo del 2018 (ver figura 7A). Para el PRI, los valores en octubre del 2017 fueron de 0.24 ± 0.004, de 0.28 ± 0.009 en marzo del 2018 y de 0.23 ± 0.004 en mayo del 2018 (ver figura 7B). El índice WBI varió con valores de entre 2.25 ± 0.005 a 2.6 ± 0.002 a través de los siete meses de muestreo, sin un patrón claro entre estaciones del año (ver figura 7C).
Discusión
Comparación diurna y nocturna de los valores del Ψ y la gs
Los valores reportados aquí del potencial hídrico y conductancia de los estomas indican que el Ψ nocturno es un mejor indicador de la variación en el estado hídrico de las plantas que el Ψ diurno. Durante el día, el Ψ no tuvo tanta variación entre estaciones del año en comparación con la noche, además de observarse una consistente correlación con la cantidad de irradiancia cotidiana, medida a partir del flujo de fotones fotosintéticamente disponibles (ver cuadro 1). En la literatura especializada hay controversias respecto a qué hora del día es la mejor para la cuantificación de estos parámetros, donde algunos autores indican que el Ψ se debe cuantificar preamanecer (Williams y Araujo 2002) y otros indican que debe realizarse la medición a medio día (Acevedo-Opazo et al., 2010) e, incluso, otros estudios no encuentran variación en estos indicadores entre tratamientos de irrigación (Mirás-Avalos et al., 2014), pero dado que en nuestro caso observamos que solo el Ψ nocturno se modificaba en función de la irrigación, sugerimos su uso como indicador.
De manera adicional, no encontramos asociación entre el contenido de agua del suelo y el Ψ, lo cual puede indicar que la disponibilidad del vital líquido en el sustrato fue suficientemente alta para evitar cualquier fluctuación, excepto durante el periodo en el que se suspendió por completo la irrigación. Este resultado puede implicar que el viñedo estudiado fue sobreirrigado, pues en ningún momento se disminuyó el Ψ por debajo de -0.5 MPa durante los veranos de estudio (fructificación, envero y postenvero), pero sí bajó durante el otoño (senescencia), caso equivalente al encontrado en otros viñedos del mundo (Mirás-Avalos et al., 2014).
Aún con el régimen de riego, nuestros valores para el potencial hídrico en el punto de marchitamiento permanente son más bajos que otros reportes, pues mientras que nosotros encontramos datos de -2.69 ± 0.12 MPa, otros estudios dan valores de -2.11 a -2.53 MPa con déficit de irrigación en Cataluña (Alsina et al., 2007), -1.08 a -1.31 MPa en invernaderos de Italia (Hochberg et al., 2017), -1.16 a -1.38 MPa en dos cultivares de Italia (Tombesi et al., 2015) y de -1.5 a -2.7 MPa en los de Hungría (Villangó et al., 2013). En las plantas de nuestro estudio, los valores del Ψ siempre fueron más positivos que el potencial en el punto de marchitamiento (figura 5). Esto implica que las plantas en el viñedo estudiado tienen la capacidad para aclimatarse efectivamente a la restricción de irrigación. Esta sería una estrategia adecuada para reducir el volumen de esta sin menoscabo de productividad, con beneficios adicionales como reducción en la incidencia de enfermedades (da Silva et al., 2018). De manera adicional, Vitis vinifera var. Cabernet Franc tiene reportada capacidad de ajuste osmótico, que permite incrementar la concentración de solutos compatibles (como almidón y prolina) en las células vegetales, promoviendo la absorción de agua en condiciones de sequía, lo que la haría una variedad apropiada para la irrigación deficitaria (Düring, 1984; Patakas y Noitsakis, 1999).
Los valores de la gs están acoplados con los del Ψ (ver figura 6), un resultado previamente encontrado en la literatura (Williams y Araujo, 2002) pero, a diferencia de investigaciones previas que cuantifican las relaciones hídricas de uvas antes del amanecer y medio día (Mullins et al., 1992), nuestro periodo de trabajo abarcó todas las horas nocturnas para el estudio de ciclos diarios. Con base en estos resultados, dada la homogeneidad en los valores del potencial nocturno respecto al diurno, podemos sugerir que las horas más apropiadas para medición de los parámetros hidráulicos en plantas es posterior al anochecer, pues logísticamente es menos demandante que la medición en madrugada, que es la convencional sugerida en la literatura. En particular, el Ψ nocturno puede ser utilizado como un indicador en esquemas de irrigación deficitaria en plantas de vid en Baja California.
Trayectorias de índices de vegetación medidos en hoja
El estudio de la variación temporal de los valores de estos índices foliares en uva es todavía muy limitado y no hay otras investigaciones para comparar las trayectorias de cambios de coloración a escala planta fuera de nuestros resultados. Es, sin embargo, posible comparar los datos que obtuvimos con evaluaciones realizadas con sensores remotos. Pero, debe aclararse que el espectro detectado por un sensor es la sumatoria de todas las reflectancias del terreno y es dependiente de la escala espacial y radiométrica. El equipo que utilizamos es un UNISPEC (SC, PPSystems), que solamente cuantifica en un cm2 de hoja el espectro completo de absorción y reflexión de la luz, con una muy alta resolución espectral, pero sin la espacial, como lo haría un sensor NDVI sobre una plataforma remota.
Nuestros datos variaron en NDVI desde 0.23 hasta 0.56, dependiendo de la expansión foliar, encontrándose valores más altos durante mayo (floración) y desde la expansión foliar en enero y febrero y bajando progresivamente durante el proceso de senescencia, desde octubre a diciembre (ver figuras 7). Otros trabajos han encontrado valores desde 0.1 hasta 0.6 para diferentes partes del mundo (Johnson, 2003; Kazmierski et al., 2011; Junges et al., 2019), pero como esos estudios han sido realizados por medio de sensores remotos, los valores de NDVI deben estar muy influenciados por las condiciones propias del terreno, incluyendo el tipo de sustrato, la densidad de plantas por hectárea y el índice de área foliar por planta, entre otros. Desafortunadamente, no encontramos valores de referencia para esta especie en relación con los índices más recientes ecofisiológicos, el PRI y el WBI. Es preciso diseñar investigaciones que permitan entender la asociación de las mediciones espectrales en planta hasta las proporcionadas por diferentes tipos de sensores remotos.
Conclusiones
Los suelos del Valle de Guadalupe difieren en propiedades físico-químicas, con los más ácidos, con mayor contenido de materia orgánica en la región sur y con los de mayor contenido de sólidos disueltos en el norte.
En el viñedo estudiado, el Ψ nocturno disminuyó durante el otoño (senescencia foliar) cuando se suspendió la irrigación, mientras que el diurno no presentó cambios entre estaciones del año (verano, durante fructificación vs. otoño, durante senescencia).
Los valores del Ψ estuvieron altamente correlacionados con la irradiación y prácticamente no hubo correlación con el contenido de agua del suelo.
El potencial hídrico en el punto de marchitamiento (-2.6 MPa) fue de los más bajos reportados en la literatura, lo cual puede indicar que las vides estudiadas pueden ser candidatas para irrigación deficitaria.
Los valores del Índice de Diferencia Normalizada de Vegetación fueron menores durante la senescencia (0.23) y mayores desde la expansión foliar a la floración (0.56).
Recomendaciones
- Cuantificar el cambio de uso de suelo y vegetación en el Valle de Guadalupe para ver si los suelos más fértiles encontrados al sur de esta región son producto de la conversión reciente a agroecosistemas.
- Evaluar esquemas de irrigación deficitaria basados en el Ψ e irrigación vespertina-nocturna en el VG y su impacto en productividad y calidad.
- Estudiar la variación espectral en hojas de vid en el Valle durante toda la temporada fenológica y su asociación con productividad y calidad.
- Cuantificar el escalamiento de la variación en índices de diferencia normalizada de vegetación a nivel hoja, sensores NDVI terrestres, sensores acoplados a vehículos aéreos no tripulados y a plataformas satelitales para detectar el nivel de exactitud de cada uno en la predicción de productividad.
_____________
Fuentes
Acevedo-Opazo, C., S. Ortega-Farias y S. Fuentes. “Effects of grapevine (Vitis vinifera L.) water status on water consumption, vegetative growth and grape quality: An irrigation scheduling application to achieve regulated deficit irrigation”, en: Agricultural Water Management. 97, vol. 7, pp. 956-964, 2010.
Alsina, M., F. De Herralde, X. Aranda, R. Save y C. Biel. “Water relations and vulnerability to embolism are not related: experiments with eight grapevine cultivars”, en: VITIS. 46, vol. 1, pp. 1-6, 2007.
Améglio, T., P. Archer, M. Cohen, C. Valancogne, F. A. Daudet, S. Dayau y P. Cruiziat. “Significance and limits in the use of predawn leaf water potential for tree irrigation”, en: Plant and Soil. 207, vol. 2, pp. 155-167, 1999.
Bartlett, M. K., C. Scoffoni, y L. Sack. “The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: a global meta‐analysis”, en: Ecology Letters. 15, vol. 5, pp. 393-405, 2012a.
Bartlett, M. K., C. Scoffoni, R. Ardy, Y. Zhang, S. Sun, K. Cao y L. Sack. “Rapid determination of comparative drought tolerance traits: using an osmometer to predict turgor loss point”, en: Methods in Ecology and Evolution. 3, vol. 5, pp. 880-888, 2012b.
Blackburn, G. A. “Spectral indices for estimating photosynthetic pigment concentrations: a test using senescent tree leaves”, en: International Journal of Remote Sensing. 19, vol. 4, pp. 657-675, 1998.
Bouyoucos, G. J. “Hydrometer method improved for making particle size analyses of soils 1”, en: Agronomy Journal. 54, vol. 5, pp. 464-465, 1962.
Boyer, J. S. Measuring the water status of plants and soils. Cambridge, Massachusetts, E.U.A., Academic Press, Inc., 1995.
Carter, G. A. y A. K. Knapp. “Leaf optical properties in higher plants: linking spectral characteristics to stress and chlorophyll concentration”, en: American Journal of Botany. 88, vol. 4, pp. 677-684, 2001.
Cavender‐Bares, J., J. A. Gamon, S. E. Hobbie, M. D. Madritch, J. E. Meireles, A. K. Schweiger y P. A. Townsend. “Harnessing plant spectra to integrate the biodiversity sciences across biological and spatial scales”, en: American Journal of Botany. 104, vol. 7, pp. 966-969, 2017.
Claudio, H. C., Y. Cheng, D. A. Fuentes, J. A. Gamon, H. Luo, W. Oechel, y D. A. Sims. “Monitoring drought effects on vegetation water content and fluxes in chaparral with the 970 nm water band index”, en: Remote Sensing of Environment. 103, vol. 3, pp. 304-311, 2006.
Cochard, H. “The basics of plant hydraulics”, en: Journal of Plant Hydraulics. INRA Editions, 2014, 1, e0001 (DE) https://bit.ly/3sOOxGA
Daesslé, L. W., L. G. Mendoza-Espinosa, V. F. Camacho-Ibar, W. Rozier, O. Morton, L. Van Dorst y A. Rodríguez-Pinal. “The hydrogeochemistry of a heavily used aquifer in the Mexican wine-producing Guadalupe Valley, Baja California”, en: Environmental Geology. 51, pp. 151-159, 2006.
Dai, A. “Drought under global warming: a review”, en: Wiley Interdisciplinary Reviews: Climate Change. 2, vol. 1, pp. 45-65, 2011.
_______ “Increasing drought under global warming in observations and models”, en: Nature Climate Change. 3, vol. 1, pp. 52-58, 2013.
Da Silva, J. R., W. P. Rodrigues, L. S. Ferreira, W. de Paula Bernado, J. S. Paixão, A. E. Patterson y E. Campostrini. “Deficit irrigation and transparent plastic covers can save water and improve grapevine cultivation in the tropics”, en: Agricultural Water Management. 202, pp. 66-80, 2018.
Del-Toro-Guerrero, F. J., T. Kretzschmar y S. H. Bullock. “Precipitation and topography modulate vegetation greenness in the mountains of Baja California, México” en: International Journal of Biometeorology. 63, vol. 10, pp. 1425-1435, 2019.
Dixon, H. H. y J. Joly. “XII. On the ascent of sap”, en: Philosophical Transactions of the Royal Society of London (B.). 186, pp. 563-576, 1895.
Düring, H. “Evidence for osmotic adjustment to drought in grapevines (Vitis vinifera L.)”, en: VITIS-Journal of Grapevine Research. 23 vol. 1, pp. 1-10, 1984.
Elmore, A. J., J. F. Mustard, S. J. Manning y D. B. Lobell. “Quantifying vegetation change in semiarid environments: precision and accuracy of spectral mixture analysis and the normalized difference vegetation index”, en: Remote Sensing of Environment. 73, vol. 1, pp. 87-102, 2000.
Espejel, I., D. W. Fischer, A. Hinojosa, C. Garcı́a y C. Leyva. “Land-use planning for the Guadalupe Valley, Baja California, Mexico”, en: Landscape and Urban Planning. 45, vol. 4, pp. 219-232, 1999.
Evert, R. F. Esau's plant anatomy: meristems, cells, and tissues of the plant body: their structure, function, and development. Nueva Jersey, E.U.A., John Wiley & Sons, 2006.
Gamon, J., L. Serrano y J. S. Surfus. “The photochemical reflectance index: an optical indicator of photosynthetic radiation use efficiency across species, functional types, and nutrient levels”, en: Oecologia. 112, vol. 4, pp. 492-501. 1997,
Garbulsky, M. F., J. Peñuelas, D. Papale y I. Filella. “Remote estimation of carbon dioxide uptake by a Mediterranean forest”, en: Global Change Biology. 14, vol. 12, pp. 2860-2867, 2008.
Hochberg, U., A. G. Bonel, R. David-Schwartz, A. Degu, A. Fait, H. Cochard y J. C. Herrera. “Grapevine acclimation to water deficit: the adjustment of stomatal and hydraulic conductance differs from petiole embolism vulnerability”, en: Planta. 245, vol. 6, pp. 1091-1104, 2017.
Intergovernmental Panel on Climate Change (IPCC). Global Warming of 1.5 °C: An IPCC Special Report on the Impacts of Global Warming of 1.5 °C Above Pre-industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty. Intergovernmental Panel on Climate Change, 2018.
Jacobsen, A. L., R. B. Pratt, S. D. Davis y F. W. Ewers. “Comparative community physiology: nonconvergence in water relations among three semi‐arid shrub communities”, en: New Phytologist. 180, vol. 1, pp. 100-113, 2008.
Johnson, L. F. “Temporal stability of an NDVI‐LAI relationship in a Napa Valley vineyard”, en: Australian Journal of Grape and Wine Research. 9, vol. 2, pp. 96-101, 2003.
Junges, A. H., D. C. Fontana y C. S. Lampugnani. “Relationship between the normalized difference vegetation index and leaf area in vineyards”, en: Bragantia. 78, vol. 2, pp. 297-305, 2019.
Kazmierski, M., P. Glémas, J. Rousseau y B. Tisseyre. “Temporal stability of within-field patterns of NDVI in non-irrigated Mediterranean vineyards”, en: Oeno One. 45(2), pp. 61-73, 2011.
McDowell, N., W. T. Pockman, C. D. Allen, D. D. Breshears, N. Cobb, T. Kolb y E. A. Yepez. “Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought?”, en: New Phytologist. 178, vol. 4, pp. 719-739, 2008.
McDowell, N. G. “Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality”, en: Plant Physiology. 155, vol. 3, pp. 1051-1059, 2011.
Meireles, J. E., B. O’Meara y J. Cavender-Bares. “Linking Leaf Spectra to the Plant Tree of Life”, en: Remote Sensing of Plant Biodiversity. Springer, pp. 155-172, 2020.
Mirás-Avalos, J. M., E. Trigo-Córdoba y Y. Bouzas-Cid. “Does predawn water potential discern between irrigation treatments in Galician white grapevine cultivars?”, en: OENO One. 48, vol. 2, pp. 123-127, 2014.
Mullins, M. G., A. Bouquet y L. E. Williams. Biology of the grapevine. Cambridge University Press, 1992.
Myneni, R. B., S. Maggion, J. Iaquinta, J. L. Privette, N. Gobron, B. Pinty y L. D. Williams. “Optical remote sensing of vegetation: modeling, caveats, and algorithms”, en: Remote Sensing of Environment. 51, vol. 1, 169-188, 1995.
Nobel, P. S. Physicochemical & environmental plant physiology. Academic press, 1999.
Patakas, A. y B. Noitsakis. “Osmotic adjustment and partitioning of turgor responses to drought in grapevines leaves”, en: American Journal of Enology and Viticulture. 50(1), pp. 76-80, 1999.
Patakas, A., B. Noitsakis y A. Chouzouri. “Optimization of irrigation water use in grapevines using the relationship between transpiration and plant water status”, en: Agriculture, Ecosystems & Environment. 106, vols. 2-3, pp. 253-259, 2005.
Peñuelas, J., I. Filella, C. Biel, L. Serrano y R. Save. “The reflectance at the 950-970 nm region as an indicator of plant water status”, en: International Journal of Remote Sensing. 14, vol. 10, pp. 1887-1905, 1993.
Pérez-Harguindenguy et al. “New handbook for standardised measurement of plantfunctional traits worldwide”, en: Australian Journal of Botany. 61, pp. 167-234, 2013.
Pettorelli, N., J. O. Vik, A. Mysterud, J. M. Gaillard, C. J. Tucker y N. C. Stenseth. “Using the satellite-derived NDVI to assess ecological responses to environmental change”, en: Trends in Ecology & Evolution. 20, vol. 9, pp. 503-510, 2005.
Romero-Toledo, Y. Variación de la edafología y estrés hídrico en Vitis vinifera L. con relación al relieve en un viñedo del Valle de Guadalupe, BC, México. Tesis de Maestría en Ciencias de la Tierra. Centro de Investigación Científica y de Educación Superior de Ensenada, 2017.
Rundel, P. W., M. T. Arroyo, R. M. Cowling, J. E. Keeley, B. B. Lamont y P. Vargas. “Mediterranean biomes: evolution of their vegetation, floras, and climate”, en: Annual Review of Ecology, Evolution, and Systematics. 47, pp. 383-407, 2016.
Schmidt, H. y A. Karnieli. “Remote sensing of the seasonal variability of vegetation in a semi-arid environment”, en: Journal of Arid Environments. 45, vol. 1, pp. 43-59, 2000.
Scholander, P. F., E. D. Bradstreet, E. A. Hemmingsen y H. T. Hammel. “Sap pressure in vascular plants: negative hydrostatic pressure can be measured in plants”, en: Science. 148, vol. 3668, pp. 339-346, 1965.
Secretaría de Fomento Agropecuario, BC, 2015 (DE) https://bit.ly/3ynH7v2
Shackel, K. A., H. Ahmadi, W. Biasi, R. Buchner, D. Goldhamer, S. Gurusinghe y G. McGourty. “Plant water status as an index of irrigation need in deciduous fruit trees”, en: HortTechnology. 7, vol. 1, pp. 23-29, 1997.
Sperry, J. S. y M. T. Tyree. “Mechanism of water stress-induced xylem embolism”, en: Plant Physiology. 88, vol. 3, pp. 581-587, 1988.
Sperry, J. S., V. Stiller y U. G. Hacke. “Xylem hydraulics and the soil–plant–atmosphere continuum”, en: Agronomy Journal. 95, vol. 6, pp. 1362-1370, 2003.
Taiz, L. y E. Zeiger. “Water balance of plants”, en: Plant Physiology. Fifth Ed. Sunderland, Massachusetts, USA, Sinauer Associates, pp. 85-105, 2010.
Tombesi, S., A. Nardini, T. Frioni, M. Soccolini, C. Zadra, D. Farinelli y A. Palliotti. “Stomatal closure is induced by hydraulic signals and maintained by ABA in drought-stressed grapevine”, en: Scientific Reports. 5, pp. 124-149, 2015.
Trenberth, K. E., A. Dai, G. Van Der Schrier, P. D. Jones, J. Barichivich, K. R. Briffa y J. Sheffield. “Global warming and changes in drought”, en: Nature Climate Change. 4, vol. 1, pp. 17-22, 2014.
Tucker, C. J. “Red and photographic infrared linear combinations for monitoring vegetation”, en: Remote Sensing of Environment. 8, vol. 2, pp. 127-150, 1979.
Tyree, M. T. y J. S. Sperry. “Vulnerability of xylem to cavitation and embolism”, en: Annual Review of Plant Biology. 40, vol. 1, pp. 19-36, 1989.
Tyree, M. T. y F. W. Ewers. “The hydraulic architecture of trees and other woody plants”, en: New Phytologist. 119, vol. 3, pp. 345-360, 1991.
Tyree, M. T. y M. H. Zimmermann. Xylem structure and the ascent of sap. Springer Science & Business Media, 2013.
Villangó, S., Z. Zsófi y B. Bálo. “Pressure-volume analysis of two grapevine cultivars ('Kékfrankos' and 'Portugieser', Vitis vinifera L.): water deficit, osmotic conditions and their possible relations with drought tolerance”, en: Vitis. 52, vol. 4, pp. 205-206, 2013.
Williams, L. E. y F. J. Araujo. “Correlations among predawn leaf, midday leaf, and midday stem water potential and their correlations with other measures of soil and plant water status in Vitis vinifera”, en: Journal of the American Society for Horticultural Science. 127, vol. 3, pp. 448-454, 2002.


